

RESEARCH
根粒共生について
マメ科などの一部の植物は、根に根粒と呼ばれる器官を形成し、根粒を介して窒素固定細菌の根粒菌と共生関係を築くことができます。この根粒共生により、植物は根粒菌から窒素栄養を受け取る一方、根粒菌には光合成産物をエネルギー源として提供します。根粒共生の成立には、根への感染、根粒形成の開始、根粒の発達、窒素固定の駆動など、複数の過程が連続して進行することが必要で、これらは植物と根粒菌の双方によって遺伝的に制御されています。2000年頃から根粒共生の制御に関わる多くの遺伝子が同定され、共生の仕組みが理解されつつありますが、未解明な点も多く、また、研究の進展による新たな疑問も生まれ続けています。
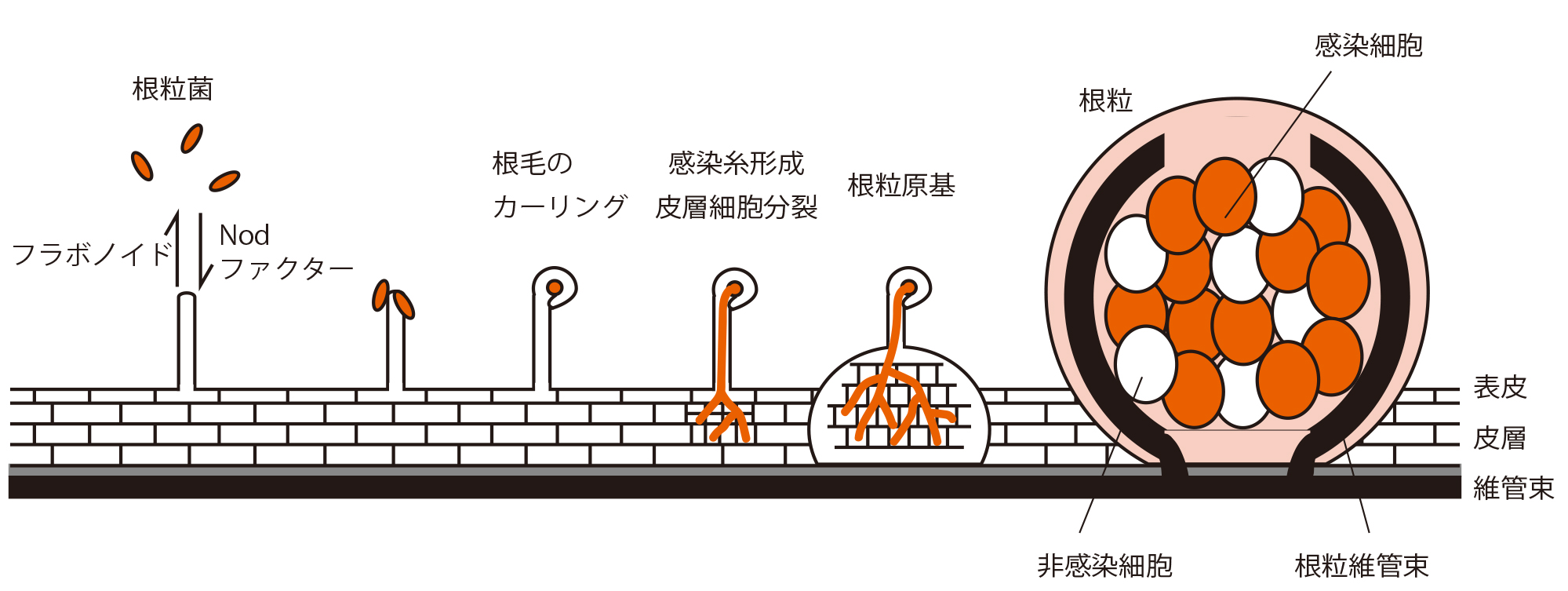
根粒共生は、植物と根粒菌の間のシグナル分子のやりとりから始まります。その後、根粒菌の感染・根への侵入と根粒器官形成が同調的に進行します。根粒の中の細胞にとりこまれた根粒菌はニトロゲナーゼを働かせて窒素固定を行います。
研究概要
環境変化に応じた根粒共生の制御機構の解明
植物に窒素栄養の供給という恩恵をもたらす根粒共生ですが、環境変化の影響を受けやすいという特徴をもっています。私たちは、根粒共生に影響を与える環境要因の一つの窒素に着目し、窒素環境に応じた根粒共生の制御の仕組みの解明に取り組んでいます。
土壌中に硝酸態の窒素栄養が十分に存在する場合、植物は根粒共生を一時的に抑制します。この制御の仕組みを解明するため、私たちはマメ科モデル植物ミヤコグサ(Lotus japonicus)を用いて研究を進めてきました。最初に、硝酸が豊富な環境でも根粒共生を行うEMS突然変異体をスクリーニングし、nitrate unresponsive symbiosis (nrsym)と名付けた複数の系統を単離しました。
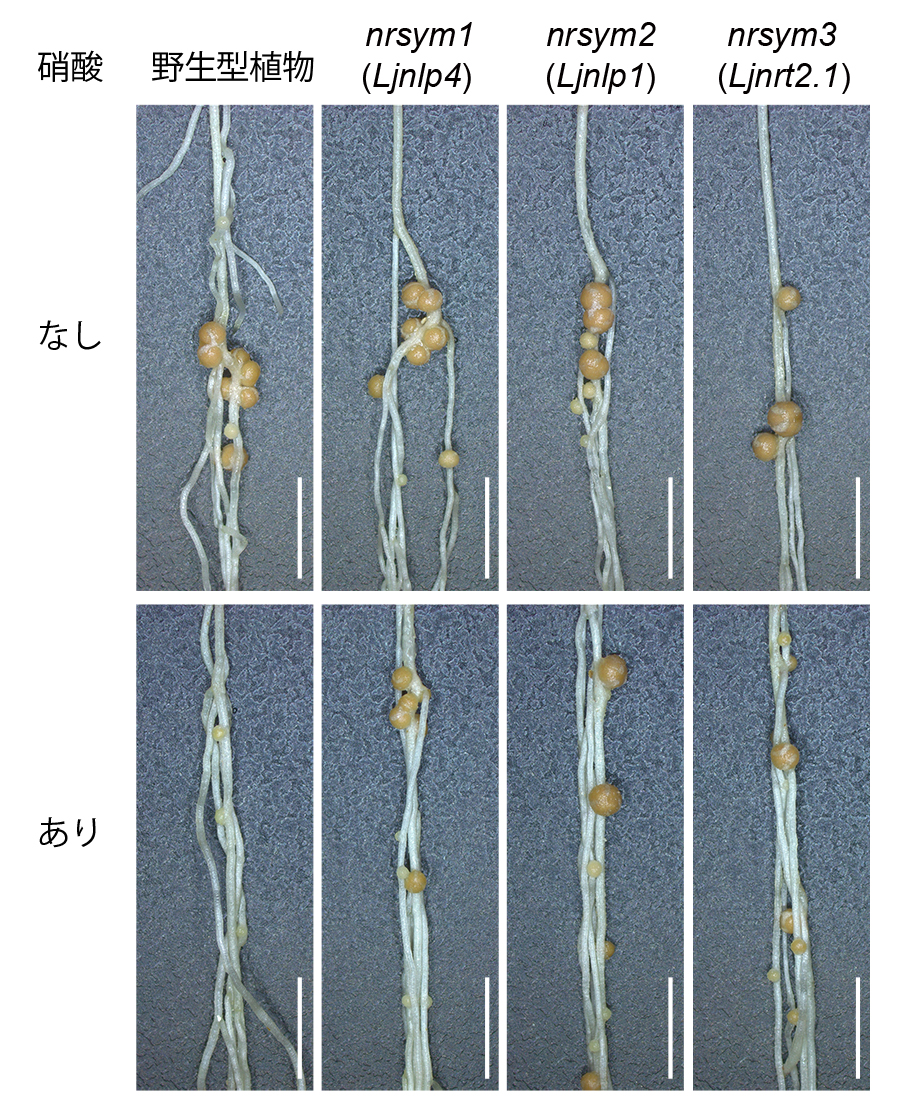
ミヤコグサのnrsym変異体は高濃度の硝酸栄養が存在する条件下でも根粒を形成します。スケールバー:5 mm
NRSYM1遺伝子はNIN-LIKE PROTEIN (NLP)転写因子の一つのLjNLP4をコードし、硝酸に応答して核へ移行して遺伝子発現を制御します(Nature Commun 2018)。また、LjNLP4がLjNIN転写因子とヘテロ二量体を形成し、根粒形成に関わる遺伝子の発現を抑制する仕組みも明らかになりました(Plant Cell 2021)。さらに、LjNLP1(NRSYM2)が硝酸イオン輸送体遺伝子LjNRT2.1の発現を介してLjNLP4の核局在を促進する働きをもつことも分かりました(Plant Cell 2022)。これらの研究成果は、マメ科植物に特有の窒素応答機構を分子レベルで解明したものです。
現在は、この制御系に関わる新たな因子の解析、アンモニアなど他の窒素栄養に対する応答の仕組みの解析、窒素と他の環境要因とのクロストークの仕組みの解析などを通して、環境変化に応じた根粒共生の制御の詳細を追求しています。
根粒共生の進化基盤の解明
根粒共生が可能な植物は、マメ目、バラ目、ブナ目、ウリ目に限られ、これらは窒素固定クレードと呼ばれています。植物の進化の過程で、窒素固定クレードの共通祖先において根粒共生能力を付与する分子進化が起こったと考えられています。根粒共生の進化基盤の解明は、新たな形質が進化の過程でどのように獲得されるのかを理解する上でも重要な課題です。

根粒共生を行うことができる植物は、マメ目、バラ目、ブナ目、ウリ目に限られており、窒素固定クレードを形成しています。
NIN転写因子は根粒の発生、根粒菌の感染、共生窒素固定に至る一連の過程を統括しています。植物の進化の過程で、NINは近縁の転写因子NLPから派生し、NLPとは異なる共生特異的な機能を獲得しましたが、その分子進化の仕組みは不明でした。最近、私たちはNINのDNA結合ドメイン直後に存在する15アミノ酸からなる新規配列FR(Following RWP-RK)がNINの機能に重要な役割を果たすことを明らかにしました。FRはNINの二量体形成を安定化し、DNA結合の柔軟性を高めることで根粒共生の制御に必要な遺伝子を含む広範な遺伝子の発現の制御を可能にしていました。さらに、NIN型FRは、被⼦植物と裸⼦植物が分岐する以前にすでに⼀部のNLPに存在していたことも判明しました。つまり、根粒共⽣そのものが出現する以前に、その分⼦基盤の⼀部はすでに⽤意されていたことになります。つまり、NINが根粒共⽣の誕⽣に合わせて「新しく発明された転写因⼦」ではなく、「既存の転写因⼦が持っていた分⼦特徴を再利⽤・洗練することで進化した」ことを⽰唆しています(Sci Adv 2026)。
現在は、この知見を基にして、NLPからNINが機能派生した分子進化の実体を全容解明することを目指しています。根粒共生の進化基盤の解明は、あらゆる植物種に根粒共生能力を付与する技術開発にもつながることが期待されます。
根粒への鉄供給機構の解明
根粒の中で行われる窒素固定反応は、根粒菌のニトロゲナーゼによって触媒されますが、この酵素複合体が働くためには鉄が必要です。しかし、どこから、どのように鉄が根粒へと運ばれて窒素固定のために使われるのか、その仕組みの多くは不明でした。
最近、私たちは根粒共生過程における遺伝子発現の変化を調査し、窒素状態に応じて発現するIRON MAN(IMA)と呼ばれるペプチドをコードする遺伝子を発見しました。このペプチドは50アミノ酸程度で、ミヤコグサにはLjIMA1からLjIMA8までの8つのバリアントがあります。LjIMA1とLjIMA2をノックアウトした植物では、根粒数の増加やサイズの減少が見られ、特に窒素固定活性が低下しました。このノックアウト植物の根粒共生の異常は鉄の蓄積不足によるものであることが分かりました。これらの発見から、IMAペプチドを介して根粒へと鉄が供給される仕組みが存在することが明らかになりました(Nature Commun 2024)。
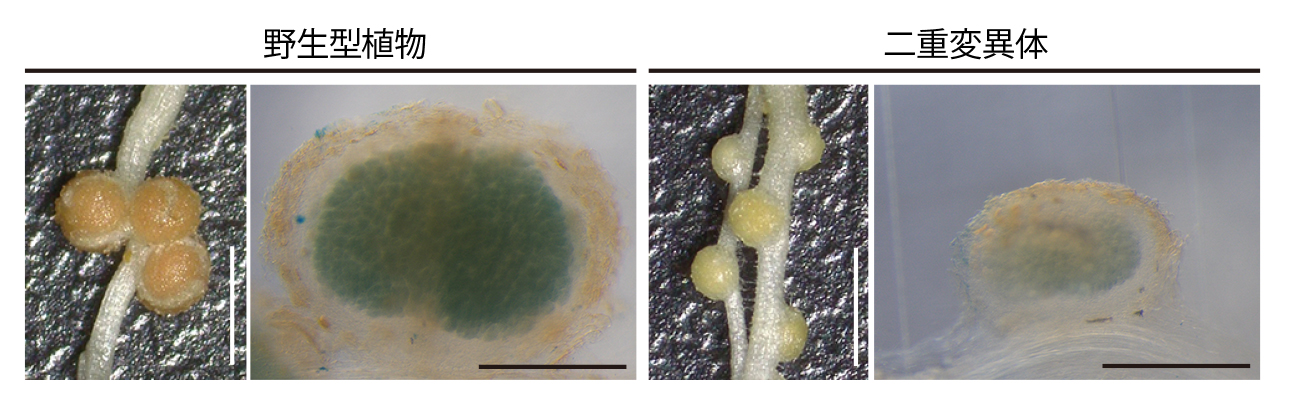
Ljima1 Ljima2二重変異体は野生型植物と比べて小さい根粒を多く形成します。根粒内部の鉄を染色すると、野生型植物の根粒は青く染色されるのに対して、二重変異体の根粒はほとんど青色に染色されないことから、鉄の存在量が少ないことが分かります。スケールバー:2 mm (白色)、500 µm (黒色)。
現在は、IMAペプチドが植物の体内を移動する仕組みや、根粒に運ばれた後の作用の仕組みに注目して解析を行っています。これらの研究の進展によって、根粒菌へと鉄が供給される仕組みの全容を理解したいと考えています。
変動環境に適応する作物の開発
20世紀初頭に開発されたハーバーボッシュ法によって合成される窒素肥料は、世界の食料生産に革命をもたらしました。しかし、この工業的窒素固定は高温・高圧で行われるためエネルギーコストが高く、温室効果ガスの5%がその影響とされています。また、先進国では過剰な窒素肥料が農地に投入されていますが、植物が吸収できる窒素量には限界があり、未使用の肥料が環境問題を引き起こしています。さらに、近未来は現在よりも高温・高CO2環境になることが予想されており、作物生産に悪影響を与える可能性が危惧されています。
私たちは、根粒菌が行う環境に優しい生物的窒素固定に着目し、このような地球規模の問題を根粒共生の力で解決したいと考えています。現在、JSTのALCA-Nextプロジェクトの支援を受け、ミヤコグサやシロイヌナズナなどのモデル植物の研究により得られた知見を活用し、将来の変動環境においても頑健に共生窒素固定を行う環境適応型のマメ科作物ダイズを開発することを目指した研究プロジェクトを進めています。